■自己組織化&自己集合
自己組織化 はみ出しコラム その1
驚くべきほど精巧な分子モーター
自己組織化によってできた細菌の小さな鞭毛の分子モーターが、どれほど複雑にできているのかを見てみよう。自己組織化の話とは少々ずれてしまうが、今後「ナノマシーン」などを実現していく上で、分子モーターの仕組みから学ぶことは決して少なくないだろう。
現時点で、分子モーターが回転する仕組みは完全に分かっているわけではないが、細胞内外の濃度差によって、細胞膜を横切る水素イオンの駆動力が関与していることは確かである。
細菌のスケールの世界では、水分子などのブラウン運動が支配的になっているが、そのランダムな力を上手に利用しているのには驚かされる。われわれが精巧な駆動機関をつくるときにまず考えることは、いかに外部からのノイズを取り除くかということにあるが、細菌たちは逆にノイズをうまく利用していることになる。このことは、ナノテクによるものづくりを今後進めていく上で、重要な示唆をしているように思われる。
また、エサなどの存在に反応して近づいたり、危険な化学物質から離れたりするときに、細菌は鞭毛の回転モードを変化させているが、そのメカニズムも驚くほどうまくできている。
大腸菌の場合、化学誘引物質が存在しているときに、鞭毛が反時計回りに回転することでプロペラの役割をして、誘引物質の方向に進む。一方で、忌避物質が存在しているときには、鞭毛を時計回りに回転させ、プロペラの機能がなくなってしばらく無秩序な動きをする。そして細菌は違う方向に動き出せるようになリ、忌避物質から逃げていく。
この二つの回転モードを切り替えている鍵は、細胞膜の特定部位に存在しているメチル基にある。化学誘引・忌避物質からでる刺激物質が、そのメチル基に付加されたり除去されたりすることで、シグナルを発生させ鞭毛の動きを制御しているのだ。
いずれナノマシーンをつくるときが来たときに、エネルギーの供給源や制御の方法などが問題となるが、自然のナノマシーンである細菌はよい見本となるだろう。
自己組織化 はみ出しコラム その2
試験管の中に現れる動物の模様いろいろ

シマウマの縞模様やキリンの斑点、それに熱帯魚のカラフルな模様を見ると、生命に特有の生き生きとした印象を受ける。しかし、こういった模様は生物だけに特有のものではなくて、試験管の中の化学反応にも現れる。そして、両者に共通するのがこの自己組織化だ。
一般によく知られている化学反応は、沈殿が生じたり、泡が発生したりするといったものだろう。ところが、あるタイプの化学反応は、一定のリズムを持って色が変化したり、縞模様が現れたりするものがある。そういった反応でとくに有名なものが「BZ反応(ベルゾーフ・ザボンチンスキ反応)」だ。例えば希硫酸中に臭素酸カリウムを加えた場合がそうだ。「自触媒作用」と呼ばれるものが、BZ反応の面白さの原因となっている。
そこで、この反応機構がどのような仕組みになっているかを少し考えてみよう。(BZ反応は非常にたくさんのステップを含んでいるので、次のモデルは化学振動に関わるステップだけを抜き出したもの。)
1. A+X→X+X
2. X+Y→Y+Y
3. Y→B
(Aは一定割合で供給される、Bは一定の濃度に保たれ残りは排出される)
ステップ1と2が自触媒反応である。確かに、1,2,3すべてを合わせて考えれば、AがBになるというだけのつまらない反応だが、振動が生じてくる理由は、その結果よりも、途中の1と2の自触媒反応の方にある。
まず、自触媒反応の特徴にしたがって、ステップ1でXは爆発的に増加する。ところが、Xが増加すると、ステップ2が起こるチャンスが高くなる。そのため、Xの増加に少し遅れて、Yが爆発的に増加する。しかし、これによってYがXを大量に使用するのでXの量が減少しステップ1の反応速度が遅くなる。そのためステップ2の反応速度も遅くなり、Yが減少する。今度はYが減ったために、Xは消費されることなく再び怒涛のように増加するチャンスがあるといったように、XとYの周期的な濃度変化の追いかけっこが続く。(実際にグラフを描くには、反応機構から微分方程式を立てて数値的に解を求める。下図)
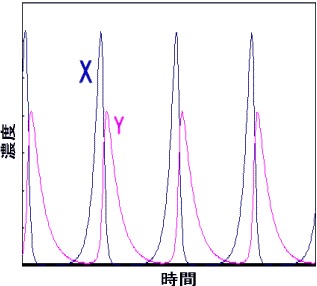
BZ反応での成分X,Yの濃度と時間の関係 |
したがって、XとYに別々の色がついていれば、周期的に色の変化が見られるというわけだ。このとき、試験管内の溶液をかき混ぜて均一にしていれば単なる色の変化が周期的に繰り返す現象になるし、溶液を混ぜずに不均一の状態にしておけば濃度差によって縞模様などが現れる。
もちろん、このBZ反応だけでは、生物の様々な模様を説明するには単純過ぎるが、他のいくつかの反応と協同作用などを考慮すれば説明できるものだと考えられている。自己触媒作用と協同作用の二つが融合した現象の一つに、レイリー・べナ-ル対流がある。この具体例には、味噌汁の中にできる模様など、身近なものがたくさんある。
自己組織化 はみ出しコラム その3
プリゴジンと非平衡熱力学・散逸構造

レイリー・べナール対流の様子。10秒という時間を連続的に撮影したもの。本来なら白い点に見えるアルミニウムの粒子が線状になっている。Photo
Copyright NOAA |
自己組織化が熱力学と深く関係していることはすぐに分かるが、いわゆるワットやカルノーたちの熱機関にはじまる古典的な熱力学や、ボルツマンなどの統計熱力学とはやや雰囲気が異なるように感じるかもしれない。
これらの熱力学は19世紀から20世紀の前半にはほとんど完成されていた。確かにこの熱力学は、科学技術の発展に多いに貢献したが限界もあった。それは、従来の熱力学の対象としている範囲が、平衡系での可逆反応と遷移であって、不可逆過程を扱うことができないことにあった。しかし、実際の世界ではほとんどが不可逆過程で、可逆過程を扱った従来の熱力学で説明できるほど単純なものではない。
こういった課題を解決するべく、外部の環境と共生して存在する「散逸構造(dissipative
structures)」の研究に大きく貢献したのが、1977年にノーベル化学賞を受賞したプリゴジン(Ilya
Prigogine)である。散逸構造では、平衡状態に近いときには秩序は薄れランダムな方向へと向かっていく一方で、平衡から大きくかけ離れたときには特定の秩序が生まれてくるという面白い性質がある。
散逸構造で最も有名な例が、先ほども挙げたべナ-ル対流だろう。これは上層の液体が下から温められたときに形成される現象だ。温度などの特定の条件が整えば対流が生じ、上層には六角形をした対流が形成される。これが形成されているのは熱が供給されている間だけで、熱が供給されなくなると消えてしまう。
プリゴジンの非平衡熱力学、散逸構造は、自己組織化とも深い関わり合いをもっている。プリゴジンの一般向けの著書は「確実性の終焉―時間と量子論、二つのパラドクスの解決(The
End of Certainty, Time, Chaos and the New
Laws of Nature)」、「複雑系の探求(Exploring
Complexity)」など。
自己組織化 はみ出しコラム その4
カオス・自己組織化・複雑系
たいていの場合自己組織化というのは、非平衡・非線形を伴い、とても複雑で、初期条件のわずかな変化によって結果が大きく左右されてしまうことがある。この点では、カオスとの関係性が考えられるわけだ。
1990年代に入って、「複雑系」というものが注目されるようになったが、これは物理学、生物学、電子工学、経済、社会学に至るまで、様々な領域でその姿を見せる横断的な学問だ。複雑系の科学の目的には、様々な学問分野で、共通の問題を同じ視点で吸えることで、背景に共通する原理を導こうとするところにある。
自己組織化やカオスは、複雑系で共通の視点を据えるための手段の一つとなっている。
|
|





